Coral
Polyp Anatomy
Home » Coral Biology » Coral Polyp Anatomy
Corals, often mistaken for plants due to their flower-like appearance,
are animals that generally exist in the form of colonies. These
colonies attach to oceanic substrate and when numerous colonies form a
consortium, a coral reef arises. Corals are members of the Animal Kingdom with
classification in the Phylum Cnidaria; further classification places them in
the class Anthozoa that is closely related to the class
Scyphyzoa (e.g. jelly fishes) and the class Hydrozoa (e.g. Hydra).
The polyp is the basic living unit of Anthozoa (further
subdivided into subclass Hexacorallia, which includes stony corals, and
subclass Octocorallia, which includes soft corals). Polyps can occur either
singularly, as in the case of anemones and some stony corals, or can form
colonies, as they do in most stony and all soft corals. A key feature of
Cnidaria is the presence of cnidocytes, or stinging cells, which the
polyp uses for prey capture and defense.
In anemones, the entire organism is comprised of a single polyp. Photo Credit: NOAA


Stony and soft corals are colonial organisms comprised of many individual polyps. Photo Credit: NOAA
 Some stony corals
can also grow as a single polyp, such as this species of Mushroom coral (Fungia
sp.)
Some stony corals
can also grow as a single polyp, such as this species of Mushroom coral (Fungia
sp.)
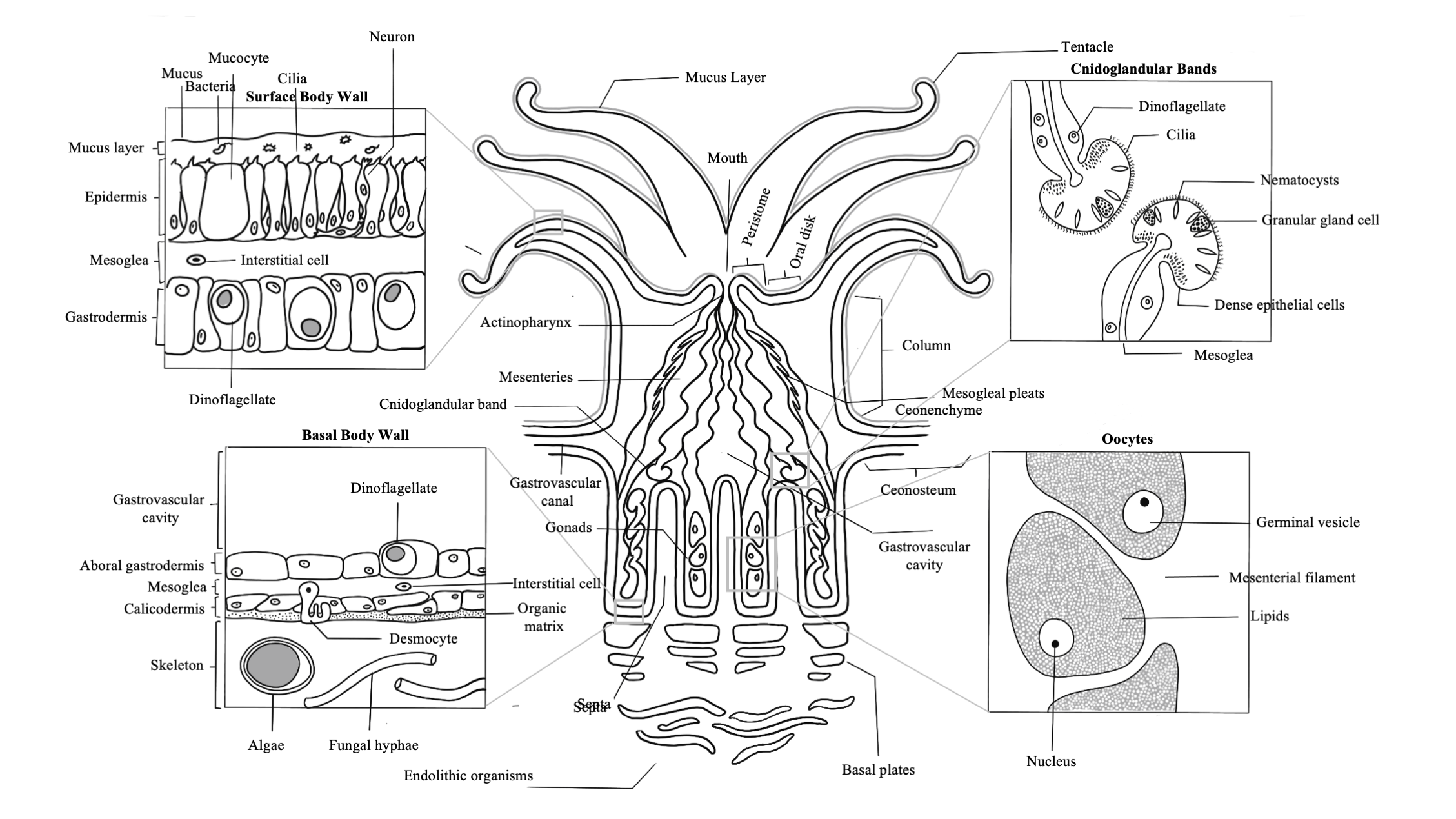
Polyp Anatomy
The fundamental structure of a coral polyp is a cylindrical sac composed
of two layers of epithelia: the surface body wall, which forms the
outermost barrier between the organism and its surroundings, and the basal
body wall, which anchors the polyp to its skeleton. Water and ingested
materials enter the polyp through the mouth at the distal (uppermost) region of
the polyp; water and waste products are expelled by the polyp through the same
opening. Respiration and excretion occur by direct exchange of molecules
through the two layers of epithelia.
 Credit: Murphy
McDonald, NOAA
Credit: Murphy
McDonald, NOAA
The portion of the polyp that is able to extend beyond the skeleton is
called the column. Located at the distal end of this region is the
polyp’s mouth, oral disk, and ring of tentacles. The mouth is
surrounded by the peristome, which is an elevated region
surrounding the mouth opening, and the oral disk, which extends
from the peristome to a ring of tentacles around its perimeter.
Tentacles are studded with stinging cells called nematocysts, and
in some cases, can be topped with a bulbous structure called an acrosphere.
Tentacles are used by the coral polyp to capture and ingest particulate
and planktonic food from the environment. For corals that live within the
photic zone (the depths where light penetrates into the ocean) the nutrients
that the coral acquires through this heterotrophic method supplements those
derived from the algal symbionts that live within its internal tissue layer,
known as the gastrodermis. The symbionts that live within corals
are microscopic dinoflagellates of the family Symbiodinacea, which
undergo photosynthesis and exchange nutrients and waste molecules with the
coral host cells. This consortium of algae within coral tissues contributes to
their color and drives the coral’s need for light in shallow reef habitats.
Single celled algae
from the family Symbiodinacea live within coral tissues and photosynthesize,
providing the coral with crucial nutrients. Photo credit: David Fleetham,
www.osf.uk.com
 Credit: Murphy
McDonald, NOAA
Credit: Murphy
McDonald, NOAA
A defining feature of anthozoans is the actinopharynx, which
is an invagination of the epidermis that forms a short muscular passageway from
the mouth to the gastric cavity. The actinopharynx controls the opening and
closing of the polyp mouth by using a ring of muscle fibers called the oral
sphincter that is located on its distal edge. The lumen of the
actinopharynx is lined with epithelial cells with elongated cilia to move
material in and out of the polyp mouth.
Extending down from the actinopharynx into the interior cavity of the
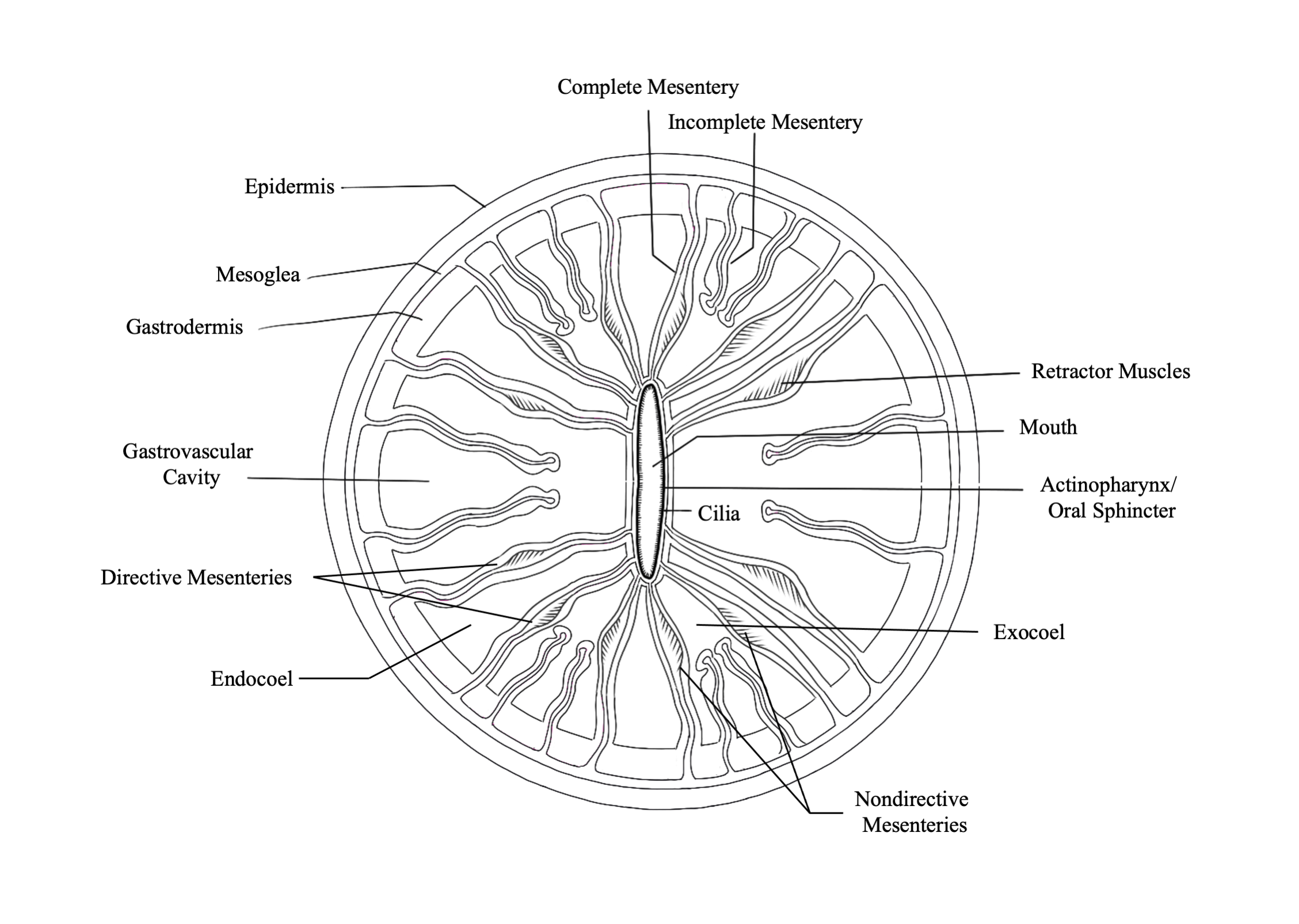
polyp are tissue partitions known as mesenteries. Mesenteries
provide structural support to the polyp column and are also important for
digestion, as they increase the surface area of the interior epithelia for gas
exchange and nutrient absorption. Retractor muscles on
mesenteries near the body wall allow the polyp to contract and extend.
Mesenteries that are attached to the actinopharynx are known as “complete” or
“perfect” mesenteries, those that are not connected are known as “incomplete”
or “imperfect.” The open space within the polyp between mesenteries is known as
the gastrovascular cavity.
Mesenterial filaments are the thickened and elongated free edges of
mesenteries below the actinopharynx. These can be stretched out of the mouth to
help capture and digest food outside and inside of the polyp using their high
abundance of granulated gland cells, which secrete digestive enzymes. Bulbous
lobes at the end of mesenterial filaments are called cnidoglandular
bands. These structures contain a battery of nematocysts for catching prey
and for defense, and of granular gland cells to aid in digestion.
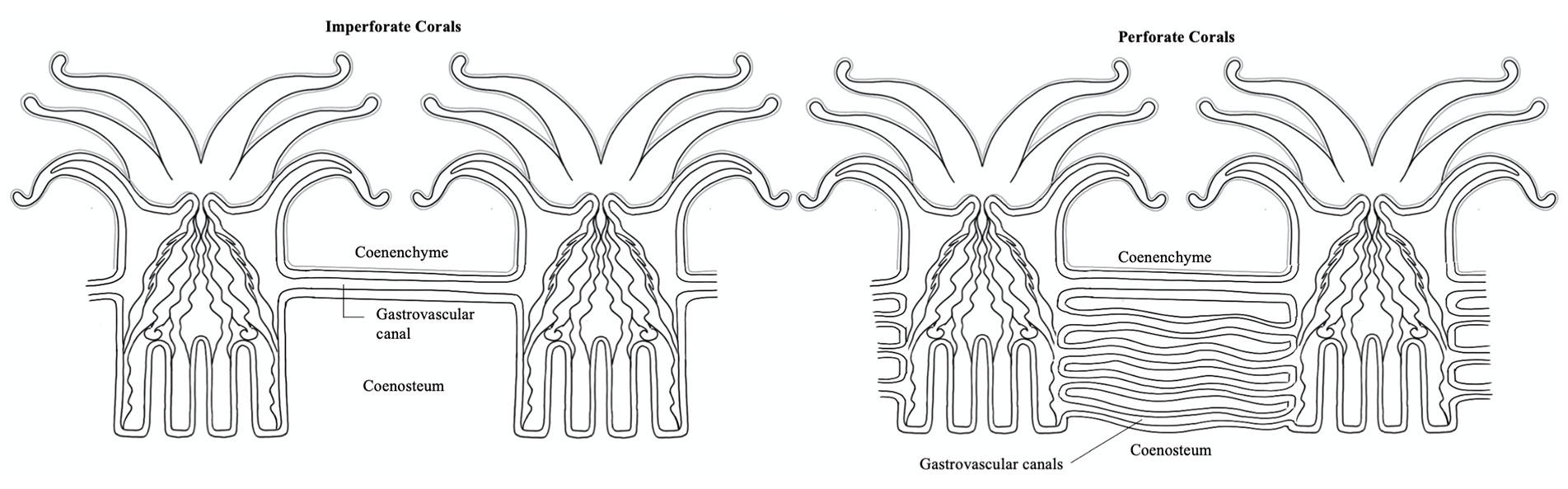
In coral colonies, the common tissue that surrounds and links polyps is
known as the coenenchyme. Within the coenenchyme, tubes
called gastrovascular canals link the gastrovascular cavities
of polyps throughout the colony. This network of tubes is what allows
individual polyps to act as a unified organism by shuttling resources and
cellular signals throughout the colony. In imperforate corals,
polyps of a colony are separated by solid skeleton (known as the coenosteum),
so this network of canals runs just under the surface of the coenenchyme above
the skeleton. In perforate corals, numerous gastrovascular canals
pierce the more porous skeleton between polyps
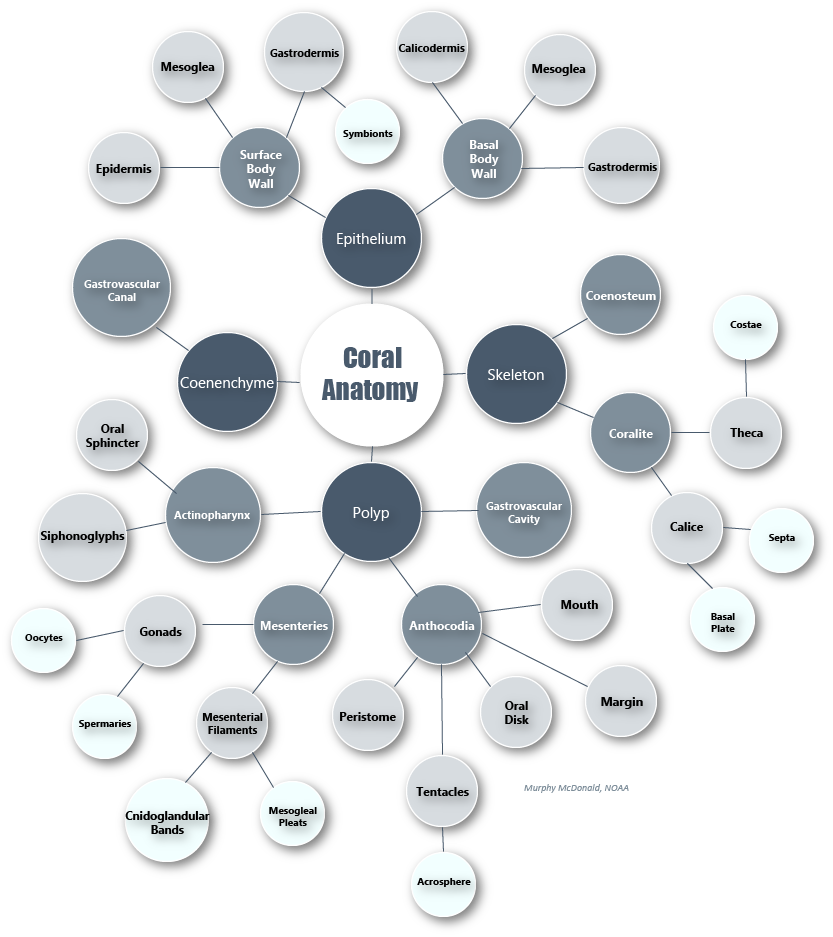
Mind map of coral polyp anatomy terms. Click to enlarge. Credit: Murphy
McDonald, NOAA
 Credit: Murphy
McDonald, NOAA
Credit: Murphy
McDonald, NOAA
Learn More about the Essentials of
Coral Biology:
Microscopic Anatomy
Learn more about the tissues that make up the coral polyp
Skeletal Anatomy
Learn about all of the components of the coral skeleton and the different
morphologies that it can display

Coral Reproduction
Learn about how corals use different reproductive modes to overcome their
immobility
![]()
Publications Relevant to Coral Anatomy
Agassiz, L. (1849) On the structure of coral animals. Proceedings of the
American Association for the Advancement of Sciences 2, 68-77.
Brown, B. E., & Bythell, J. C. (2005). Perspectives on mucus
secretion in reef corals. In Marine Ecology Progress Series (Vol. 296).
https://doi.org/10.3354/meps296291
Burton, P. M. (2008). Insights from diploblasts; the evolution of
mesoderm and muscle. In Journal of Experimental Zoology Part B: Molecular and
Developmental Evolution (Vol. 310, Issue 1).
https://doi.org/10.1002/jez.b.21150
Cairns, S. D. (1991). A revision of the Ahermatypic Scleractinia of the
Galapagos and Cocos Islands. Smithsonian Contributions to Zoology, 504.
https://doi.org/10.5479/si.00810282.504
Chornesky, E., and Peters, E. (1987) Sexual Reproduction and colony
growth in the reef coral Porites astreoides. Biological Bulletin 172, 161-177.
Epp, L., Smid, I., & Tardent, P. (1986). Synthesis of the mesoglea by
ectoderm and endoderm in reassembled hydra. Journal of Morphology, 189(3).
https://doi.org/10.1002/jmor.1051890306
Fautin, D., and Mariscal, R. (1991) Cnidaria: Anthozoa, in Microscopic
Anatomy of Invertebrates: Placozoa, Porifera, Cnidaria, and Ctenophora (eds F. harrison
and J. Westfall), Vol. 2. Wiley-Liss, New York, NY, pp. 267-358
Galloway, S., Work, T., Bochsler, V., et al. (2007) Coral Disease and
Health Workshop: Coral Histopathology II. NOAA Technical Memorandum NOS NCCOS
56 and CRCP 4. National Oceanic and Atmospheric Administration, Silver Spring,
MD.
Gateño, D., & Rinkevich, B. (2003). Coral polyp budding is probably
promoted by a canalized ratio of two morphometric fields. Marine Biology,
142(5). https://doi.org/10.1007/s00227-003-1009-8
Gladfelter, E. H. (1932) Circulation of fluids in the gastrovascular
system of the reef coral Acropora cervicornis. The Biological Bulletin, 165(3).
https://doi.org/10.2307/1541469
Goldberg, W. M. (2002). Feeding behavior, epidermal structure and mucus
cytochemistry of the scleractinian Mycetophyllia reesi, a coral without
tentacles. Tissue and Cell, 34(4).
https://doi.org/10.1016/S0040-8166(02)00009-5
Goldberg, W. M. (2002). Gastrodermal structure and feeding responses in
the scleractinian Mycetophyllia reesi, a coral with novel digestive filaments.
Tissue and Cell, 34(4). https://doi.org/10.1016/S0040-8166(02)00008-3
Goreau, T. F. (1956) A study of the Biology and Histochemistry of Corals.
Ph. D. thesis, Yale University.
Grimmelikhuijzen, C. J., & Westfall, J. A. (1995). The nervous
systems of cnidarians. In EXS (Vol. 72).
https://doi.org/10.1007/978-3-0348-9219-3_2
Lang, J. (1984) Whatever works: The variabie importance of skeletal and
of non-skeletal shcaracters in scleractinian taxonomy. Paleontographica
Americana 54, 18-44.
Lang, J. C., & Chornesky, E. (1990). Competition between
scleractinian reef corals – a review of mechanisms and effects. In Ecosystems
of the world. Volume 25: Coral reefs (Vol. 25).
Lesser, M. P., Mazel, C. H., Gorbunov, M. Y., & Falkowski, P. G. (2004).
Discovery of symbiotic nitrogen-fixing cyanobacteria in corals. Science,
305(5686). https://doi.org/10.1126/science.1099128
Marshall, A. T., & Wright, O. P. (1993). Confocal laser scanning
light microscopy of the extra-thecal epithelia of undecalcified scleractinian
corals. Cell & Tissue Research, 272(3). https://doi.org/10.1007/BF00318560
Martindale, M. Q., Pang, K., & Finnerty, J. R. (2004). Investigating
the origins of triplosblasty: “Mesodermal” gene expression in a diploblastic
animal, the sea anemone Nematostella vectensis (phylum, Cnidaria; class,
Anthozoa). Development, 131(10). https://doi.org/10.1242/dev.01119
Mullen, K., Peters, E., and harvell, C. (2004) Coral resistance to
disease, in Coral Health and Disease (eds E. Rosenberg and Y. Loya).
Springer-Verlag, Berlin, pp. 377-399.
Muscatine, L., Ferrier-Pagès, C., Blackburn, A., Gates, R. D.,
Baghdasarian, G., & Allemand, D. (1998). Cell-specific density of symbiotic
dinoflagellates in tropical anthozoans. Coral Reefs, 17(4). https://doi.org/10.1007/s003380050133
Muscatine, L., Tambutte, E., & Allemand, D. (1997). Morphology of
coral desmocytes, cells that anchor the calicoblastic epithelium to the
skeleton. Coral Reefs, 16(4). https://doi.org/10.1007/s003380050075
Nothdurft, L. D., & Webb, G. E. (2009). Clypeotheca, a new skeletal
structure in scleractinian corals: A potential stress indicator. Coral Reefs,
28(1). https://doi.org/10.1007/s00338-008-0439-7
Östman, C. (2000). A guideline to nematocyst nomenclature and classification,
and some notes on the systematic value of nematocysts. Scientia Marina,
64(SUPPLEMENT 1). https://doi.org/10.3989/scimar.2000.64s131
Palmer, C. v., Bythell, J. C., & Willis, B. L. (2010). Levels of
immunity parameters underpin bleaching and disease susceptibility of reef
corals. The FASEB Journal, 24(6). https://doi.org/10.1096/fj.09-152447
Palmer, C. v., Modi, C. K., & Mydlarz, L. D. (2009). Coral
fluorescent proteins as antioxidants. PLoS ONE, 4(10).
https://doi.org/10.1371/journal.pone.0007298
Palmer, C. v., Mydlarz, L. D., & Willis, B. L. (2008). Evidence of an
inflammatory-like response in non-normally pigmented tissues of two
scleractinian corals. Proceedings of the Royal Society B: Biological Sciences,
275(1652). https://doi.org/10.1098/rspb.2008.0335
Peters, E., (1984) A Survey of the Normal and Pathological Histology of
Scleractinian Corals with Emphasis on the Effects of Sedimentation Stress. Ph.
D. dissertation, University of Rhode Island.
Peters, E., Price, K., & J. Borsay Horowitz, D. (2005). Histological
preparation of invertebrates for evaluating contaminant effects. In Techniques
in Aquatic Toxicology, Volume 2. https://doi.org/10.1201/9780203501597.ch36
Petes, L. E., Harvell, C. D., Peters, E. C., Webb, M. A. H., &
Mullen, K. M. (2003). Pathogens compromise reproduction and induce melanization
in Caribbean sea fans. Marine Ecology Progress Series, 264. https://doi.org/10.3354/meps264167
Puverel, S., Tambutté, E., Zoccola, D., Domart-Coulon, I., Bouchot, A.,
Lotto, S., Allemand, D., & Tambutté, S. (2005). Antibodies against the
organic matrix in scleractinians: A new tool to study coral biomineralization.
Coral Reefs, 24(1). https://doi.org/10.1007/s00338-004-0456-0
Seipel, K., & Schmid, V. (2006). Mesodermal anatomies in cnidarian
polyps and medusae. In International Journal of Developmental Biology (Vol. 50,
Issue 7). https://doi.org/10.1387/ijdb.062150ks
Sharp, K. H., Ritchie, K. B., Schupp, P. J., Ritson-Williams, R., &
Paul, V. J. (2010). Bacterial acquisition in juveniles of several broadcast
spawning coral species. PLoS ONE, 5(5).
https://doi.org/10.1371/journal.pone.0010898
Vargas-Ángel, B., Peters, E. C., Kramarsky-Winter, E., Gilliam, D. S.,
& Dodge, R. E. (2007). Cellular reactions to sedimentation and temperature
stress in the Caribbean coral Montastraea cavernosa. Journal of Invertebrate
Pathology, 95(2). https://doi.org/10.1016/j.jip.2007.01.003
Veron, J. (2000) Corals of the World, Volume 1, Australian Institute of
Marine Science, Townsville, Queensland, Australia.
Zilberberg, C., & Edmunds, P. J. (1999). Patterns of skeletal structure
variability in clones of the reef coral Montastraea franksi. Bulletin of Marine
Science, 64(2).
Zilberg, C., and Edmunds, P. (1999) Patterns of skeletal structure
variability in clones of the reef coral Montastrea franksii. Bulletin of Marine
Science, 64, 373-381
Comments